
Abstract
Keywords
Introduction
Epigenetics is the study of heritable modifications in gene expression and reversible form of gene regulation that is not due to changes in the DNA sequence, it is accepted as ‘‘the assessment and study of changes in gene function that are meiotically and/or mitotically heritable without any structural changes in DNA strand. The epigenetic propose, as proposed by Conrad Waddington in the 1950s, is a branch of cell biology that evaluates the causal interactions between genes and their products, this definition points to all molecular mechanisms modulating the gene expression (1). Epigenetic alterations are crucial for the cell development and differentiation of the different cell types, as well as for normal cellular processes such as the inactivation of X-chromosome in female mammals and gene silencing. These processes can involve chemical modifications include histone acetylation, DNA methylation, ubiquitination, and phosphorylation. Recent in-vitro studies have indicated that epigenetics plays a major role in many types of cancers (2-11) such as hepatocellular carcinoma (HCC) (12-16). HCC, the predominant form of adult liver cancer, is one of the most common cancer and remains a major health problem worldwide (17). The dysregulation of the epi-genome and epigenetic control lead to aberrant gene expression, which regulates cancer induction and progression. Therefore, the role of epigenetic compounds has become significant in reverting the malignant phenotype (18, 19). Various cellular actions, comprising tumor suppressor genes (TSGs) expression, cell proliferation, DNA repair, and cell apoptosis, are regulated by acetylation and deacetylation. Histone deacetylases (HDACs) and histone acetylases (HATs) are enzymes involved in chromatin remodeling histone acetylation and deacetylation respectively. These enzymes play an important role in gene expression. HDACs and HATs are used to regulate the gene transcription by removing and/or preserving the acetyl group on specific histones. This activity can condense or relax the conformation of the DNA, changing the accessibility zones for transcription machinery. These changes result in different cell biological functions, including cell proliferation and differentiation. Reversible acetylation is one of the most post-translational modifications in eukaryotic cells. Aberrant histone protein acetylation, particularly histone deacetylation, has been shown in numerous human cancer types, dysregulation of HDACs has been implicated in the histo-pathogenesis of cancers (20, 21). Epigenetic modification by inhibiting HDAC activity is an emerging approach in cancer treatment. Histone deacetylase inhibitors (HDACIs) such as panobinostat, valproic acid (VPA), vorinostat, sodium butyrate, trichostatin A, belinostat, and romide-psin have demonstrated antitumor efficacy via activation of various molecular mechanisms. These compounds have strong anti-cancer effects, inducing cell growth arrest, cell differentiation, cell apoptosis (programmed cell death), cell invasion, and suppressing angiogenesis (22, 23). HDACIs play their apoptotic roles through multiple mechanisms such as extrinsic/cytoplasmic and intrinsic/mitochondrial molecular mechanisms (24-26). The present review highlights the role of HDACs in hepatocellular carcinoma (HCC) and the molecular mechanisms through which HDACIs play their apoptotic effects on HCC including extrinsic and intrinsic molecular mechanisms.
Epigenetic Dysregulation in Cancer induction and Development
In human cancers, common epigenetic changes include histone modification, DNA hypermethylation, and noncoding RNA dysregulation. DNA hypermethylation of CpG islands has emerged as an epigenetic mechanism of silencing TSGs. Recent evidence suggest that cancer is associated with the accumulation of cells with aberrant CpG island hypermethylation. Increasing evidence show that DNA hypermethylation of CpG islands is a common molecular mechanism in cancer induction and progression (27). In addition to DNA methylation, numerous molecular mechanisms of histone modification contribute to regulating gene expression epigenetically. Histone acetylation neutralizes the negative charge of DNA strands leads to gene transcription and expression (28). Chromatin remodeling by histone modific-ation is an example of epigenetic gene regulation. Histone deacetylases (HDACs) have recently been shown to modify a variety of other proteins that are involved in diverse cellular processes. The activity of these enzymes results in chromatin compaction, TSGs silencing, and cancer induction (29). Subsequent characterization of TSGs revealed their involvement in cancers and various mechanisms that protect animals against tumorigenesis and tumor development (30).
Histone acetylases and histone deacetylases biological activity
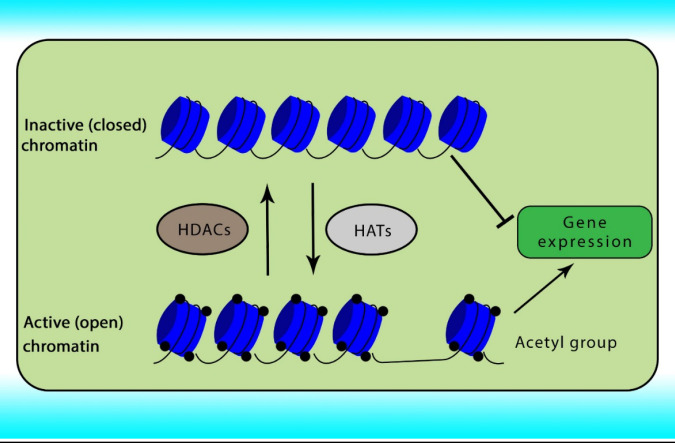
Reversible acetylation is one of the most abundant post-translational histone modifications in eukaryotic cells. Acetylation and deacetylation of histone are achieved by the opposite actions of two groups of enzymes comprising HATs and HDACs. Abnormal expression of HDACs has been indicated in various cancers. The acetylation of histone neutralizes the positive charge of the histone, relaxes the chromatin strands to facilitate the binding of transcription factors, relaxing the DNA strands and enabling greater accessibility of the transcription machinery, and subsequently gene transcription. In contrast, histone deacetylation induces chromatin compaction and gene silencing (31, 32). The role of HATs and HDACs on chromatin conformation has been shown in Figure 1.
Histone deacetylases classification
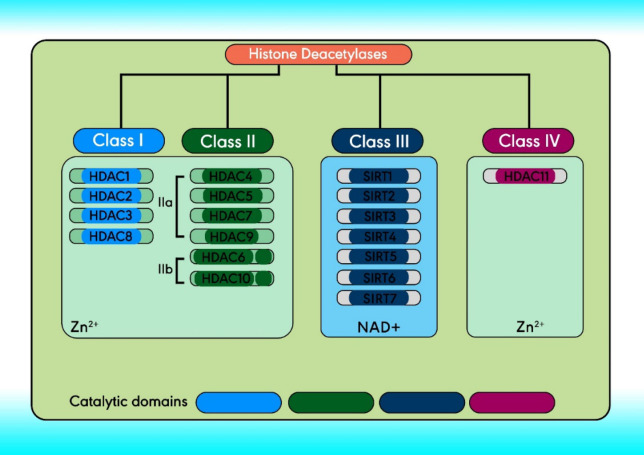
There are 18 HDACs in humans divided into four classes comprising classes I, II, and IV. Based on structure, Class III and the yeast Sir2 protein are homologous, both of which require NAD+ as a cofactor instead of Zn2+. Class III HDACs include seven mammalian sirtuins (SIRT1–7), localized in the mitochondria (SIRT3-5), nucleus (SIRT1, and SIRT6-7), and cytoplasm (SIRT2) (33-35). Class I (located within the nucleus) comprises HDACs 1- 3, and 8. Class II can travel between the cytoplasm and the nucleus and can be further subdivided, class IIa and IIb (36). The structure and classification of HDACs are indicated in Figure 2 (37).
Epigenetic dysregulation in hepatocellular carcinoma
HCC is characterized by the presence of epigenetic changes, including post-translational modifications of histone and promoter DNA hypermethylation, which affect the expression of numerous genes critical for cancer induction and progression. Strong data suggest that histone deacetylation plays an important role in HCC (38). It is now possible to analyze epigenetic abnormalities associated with various cancers (e.g. HCC). Various histone modifications are considered with transcriptional regulatory mechanisms associated with gene expression changes in HCC (39).
Histone deacetylases in HCC
Recent works on TSGs suggest that HDACs are involved in tumor formation and progression of HCC. The expression of HDACs has been linked to clinicopathological factors in HCC. The common HDACs reported in HCC have been shown in Table 1 (40-47).
Histone Deacetylase Inhibitors
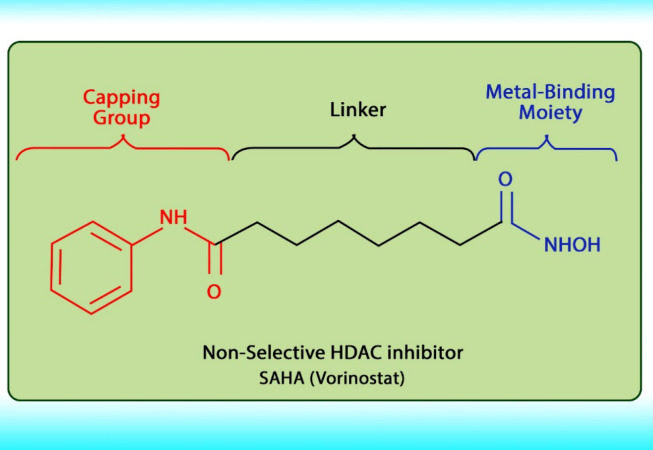
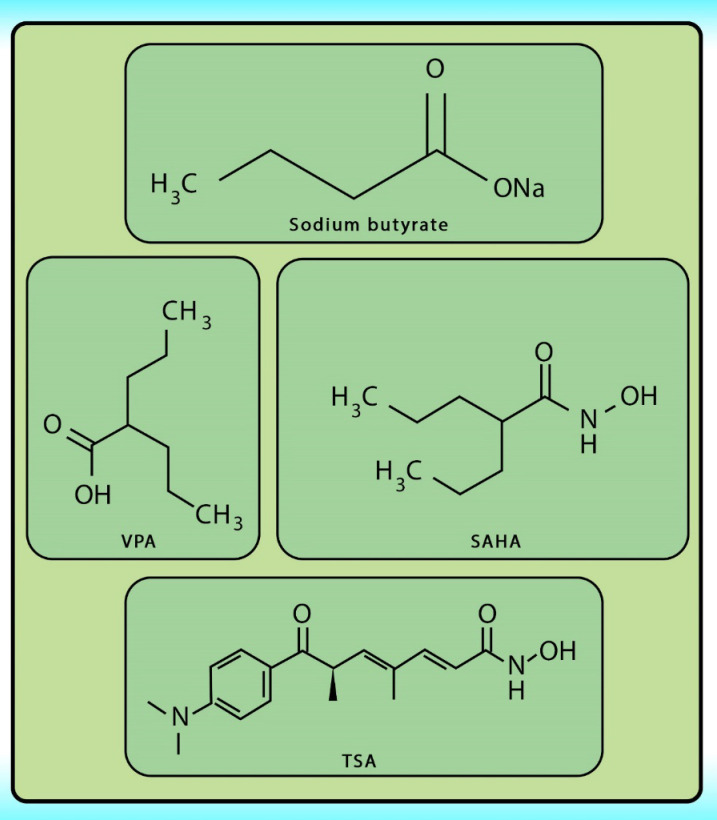
Histone acetylation has been demonstrated to be an important molecular mechanism that controls gene expression. Acetylation and deacetylation of histones cause chromatin decondensation and condensation respectively (48). Steady-state of acetylation results from the balance between the acetylation and deacetylation of histone which determines the level at which a gene is expressed (49). The changes in HAT/HADC activity balance have effect on: (1) alteration in gene expression profile as well as to the change of some signaling pathways; (2) proteasomal degradation; (3) protein kinase C activity and (4) DNA methylation status. In numerous cancers, the shift to an increased acetylation/deacetylation ratio increases chromatin structure relaxation and gene transcription. HDACIs are a diverse group of agents, which vary in biological activity, structure, and specificity. Generally, these compounds contain a capping group, a zinc-binding domain, and a straight-chain linker connecting the two parts (Figure 3). HDACIs act specifically against HDACs. Based on their chemical structures, they can be classified into five classes: (1) short-chain fatty (aliphatic) acids; (2) hydroxamic acids (hydroxamates); (3) cyclic tetrapeptides; (4) benzamides; and (5) sirtuin inhibitors (50). They may also be classified according to their specificity for HDACs. The pan‐deacetylase inhibitors comprising vorinostat, panobinostat, belinostat, trichostatin A, and resminostat. Valproate and butyrate inhibit class I and IIa HDACs, whereas entinostat, mocetinostat, and romidepsin are considered to be the class I specific. Tubacin is HDAC6 specific (51). The HDACIs classification has been indicated in Table 2 (52-54). The chemical structure of common HDACIs used in HCC has been demonstrated in Figure 4.
Target and molecular mechanisms of HDACIs in HCC
The level of histone deacetylation is associated with cancer induction and progression. HDACIs play their roles through inhibition of HDACs activity in HCC. The target of HDACIs in HCC has been shown in Table 3 (55-60) and also various molecular mechanisms by which these compounds affect HCC have been indicated in Table 4 (61-67).
Epigenetic therapy with Histone deacetylase inhibitor
There are several strategies related to epigenetic therapy in the field of cancer of which is based on the use of HDACIs. These compounds play their roles through various pathways one of which is based on selective induction of apoptosis via the intrinsic pathway and by influencing the expression of proteins such as Bcl-2, Bcl-XL, Mcl-1, and XIAP (68), as well as through activation of the DR5, DR4, and Fas (69). Additionally, HDACIs can engage the extrinsic apoptotic pathway through upregulation of DRs expression, reductions in c-FLIP, and upregulation of ligands such as TRAIL (70).
Apoptotic regulators
Over the last decade, the role of apoptosis regulators such as the BCL-2 family and their molecular mechanisms has attracted attention. This family can be classified into two groups: a group modulates mitochondrial action and another group regulates the activity of caspases. These proteins regulate the intrinsic apoptotic pathway. Several proteins of the second group, e.g. XIAP or FLIP block the activity of caspases. Cross-talk between the extrinsic and intrinsic pathways exists (71).
Apoptosis is regulated by intrinsic and extrinsic ligands. This process is controlled by the diversity cell signals pathway and is involved in the regulation of cell fate survival or death. As the cross-talk organelles, the mitochondria can connect the different apoptosis pathways (72).
The apoptotic pathways
There are two major molecular mechanisms of cell death including necrosis and apoptosis. The cells undergo necrosis because of external injury, while apoptosis is a programmed cell death because of internal or external stimuli which are controlled by numbers of complex proteins. It occurs through two main pathways comprising extrinsic and intrinsic apoptotic pathways.
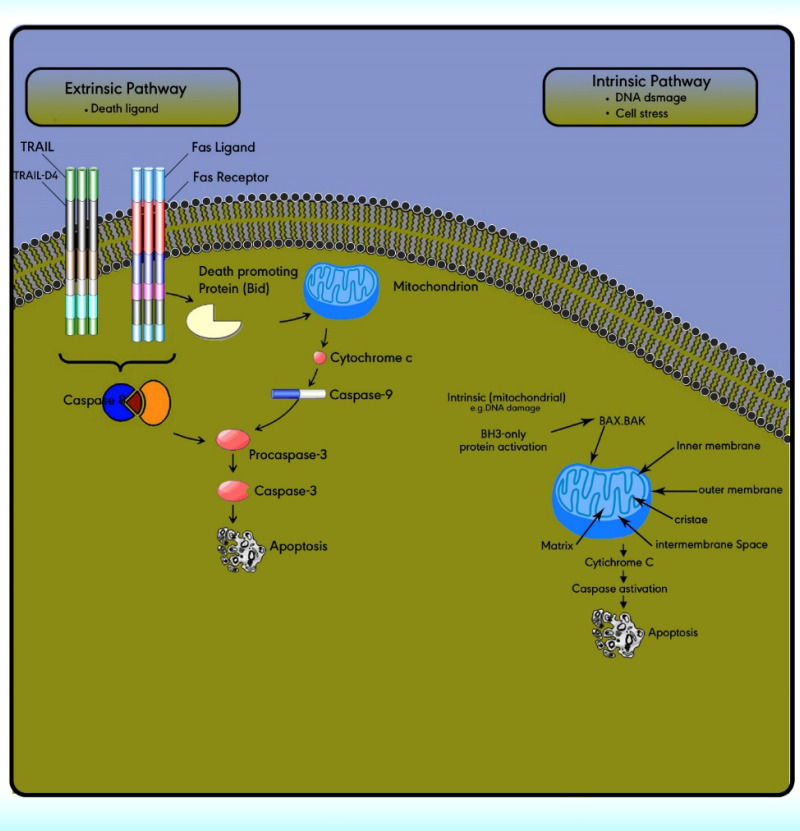
The extrinsic (death receptor) apoptotic pathway is triggered through the Fas death receptor (73). The intrinsic (mitochondrial) apoptotic pathway results in the release of cytochrome‐c from the mitochondria which activates the death signal (74). Both pathways lead to a final common pathway which activate caspases that cleave regulatory molecules, lead up to the death of the cell (Figure 5) (75).
Extrinsic pathway or death receptor pathway
The extrinsic apoptotic pathway includes numerous protein members comprising the death receptors (DRs), the Fas ligand, the Fas‐associated death domain, the Fas complexes, caspases 8, and 10 (Figure 5). The extrinsic pathway is initiated with the ligation of cell surface receptors. DRs may belong to the tumor necrosis factor family (TNF), which is composed of several members. Fas is a TNF and is also called Apo‐1 or CD95. Currently, six DRs are known, comprising TNF receptor 1, Fas, p55, p60, DR3, TNF-related apoptosis-inducing ligand receptor 1, TRAIL-R2, and DR6 (76).
The extrinsic/death receptor pathway is initiated via binding of the cell surface death receptors, DRs, with their ligands. After activation, the intracellular domains of these receptors bind to the adaptor protein Fas-associated death domain (FADD) or TNFR1-associated death domain protein (TRADD) to form the death-inducing complex with the recruitment of pro-caspase 8. Then, the activated procaspase 8 serves as the ‘initiator’ caspase, further activating downstream effectors such as caspase 7 to initiate apoptosis induction (77). In some cell types, the activation of caspase 8 may be the only requirement to execute death (78).
Intrinsic pathway or the mitochondrial pathway
The intrinsic pathway is activated via the loss of cell growth factor signals or in response to intracellular stimuli. Mitochondria have an outer membrane (OM), an inner membrane (IM), and an intermembrane space (IMS). The IMS includes several proteins involved in cell apoptosis and cell death induction, such as cytochrome c, Smac/DIABLO, Omi/HtrA2, apoptosis-inducing factors, and EndoG.
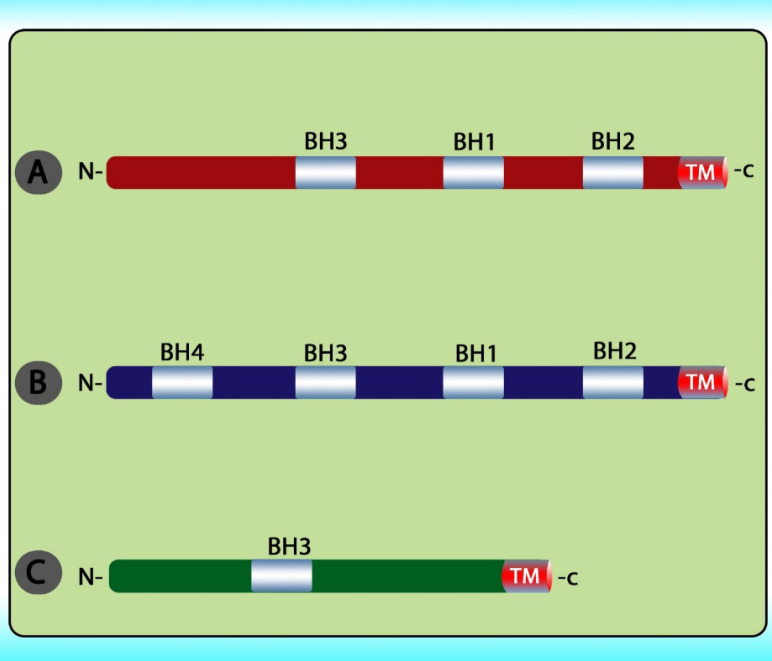
The stimuli change the permeability of the inner mitochondrial membrane and release the pro-apoptotic proteins from the intermembrane space into the cytoplasm. After the release of these proteins into the cytoplasm, cytochrome c stimulates apoptosome formation followed by caspase 9 activation. Caspase 9 activates caspases 3, 6, and 7 leads to cell death (Figure 5) (79). The intrinsic pathway is controlled through interactions between the members of the Bcl-2 protein family. This family contains 20 proteins divided into 3 groups: one anti-apoptotic group and two pro-apoptotic groups. The members of this family can be identified via Bcl-2 homology domains (BH1 to BH4). Each of the BH domains has a different action. Whereas most anti-apoptotic proteins comprise all domains and protect cells exposed to various conditions (Figure 6). These proteins are divided into two subgroups. The first group contains BH3-only domain proteins whereas the second group contains the BH 1-3 domains. Anti-apoptotic proteins exert their function by binding the member’s Bak and Bax, preventing mitochondrial damage (80). The function and classification of the BCL-2 family have been shown in Table 5 (81-88).
The common HDACs reported in HCC
| Class | Member | Localization |
|---|---|---|
| class I | HDAC 1-3, and 8 | Nucleus |
| IIa | HDAC4-7, and 9 | Nucleus/cytoplasm |
| IIb | HDAC6 and 10 | Cytoplasm |
| IV | HDAC 11 | Nucleus |
HDACIs classification
| Class | Member | Target (HDAC specifity) |
|---|---|---|
| Short-chain fatty acids | Valproic acid, | I, IIa |
| Hydroxamic acids | TSA | Pan |
| Cyclic tetrapeptides | Romidepsin | I |
| Benzamides | 4SC202 | I |
| Sirtuins inhibitors | EX-527 | SIRT 1 |
The target of HDACIs in HCC
| Class | Member | Target HDAC class | Cell line | Reference |
|---|---|---|---|---|
| short-chain fatty acids | Valproic acid (VPA), | class I, IIa | hepatocellular carcinoma HepG 2 cells | A |
| Aliphatic fatty acid | sodium butyrate | class I, IIa | SMMC-7721 and HepG2 cells | B |
| Aliphatic fatty acid | sodium butyrate | class I, IIa | HCC SMMC-7721 and HepG2 cells | C |
| Hydroxamic acid | TSA | Pan | HCC SMMC-7721 and HepG2 cells | C |
| Hydroxamic acid | trichostatinA (TSA) | Pan | Hep3B,HepG2, Hep3B | D |
| Hydroxamates | (SAHA) | Pan | Bel-7402, HepG2 | E, F |
HDACIs pathways in HCC
| Class | Member | Apoptosis pathway | Tissue or cell line | Reference |
|---|---|---|---|---|
| short-chain fatty acids | Valproic acid (VPA), | Intrinsic | HepG2, BEL-7402, and SMMC-7721 | A |
| Aliphatic fatty acid | sodium butyrate | Intrinsic | HuH-6, HepG2, HuH-6, HuH-7, Hep-G2, and PLC/PRF/5 cells | B, C |
| Hydroxamic acid | TrichostatinA (TSA) | Intrinsic | HepG2, MH1C1, Hepa1–6 and Hep1B | D |
| Hydroxamates | Suberoylanilide hydroxamic acid (SAHA) | Intrinsic | HCCLM3, 7703K, SMMC7721, BEL7402, and HepG2 cell lines | E, F, G |
The function and classification of the BCL-2 family
| Group | Member | Mechanism of action | Subcellular localization | Domain |
|---|---|---|---|---|
| Anti-apoptotic BCL-2 proteins | BCL-2 | It inhibits apoptosis by the preservation of mitochondrial membrane integrity. | (1) Nuclear envelope | Contain four BH (1-4) domains |
| BCL-W | It reduces cell apoptosis under cytotoxic conditions. | Exclusively on the mitochondrion | ||
| BCL-XL | It inhibits cytochrome c release that inhibits activation of the cytoplasmic caspase cascade by cytochrome c | The transmembrane molecule in the mitochondria | ||
| MCL1 | It interacts with Noxa, BAK1, BCL2L11, Bcl-2-associated death promoter, PCNA. | Nucleus, mitochondria | ||
| pro-apoptotic proteins | BAK | It undergoes conformational changes to form larger aggregates during apoptosis | Integral mitochondrial membrane protein | Contain three conserved BH domains |
| BAX | Release of apoptogenic factors like cytochrome c, activation of caspase cascade | Cytosol | ||
| BOK | Major functions of BOK are exerted on the ER membranes and the Golgi and that it induces apoptosis in a manner dependent on BAK and BAX. | Nucleus | ||
| pro-apoptotic BH3-only proteins | BID | Bid, free ‘activator’ type BH3 only protein, which can then activate Bak and Bax. | Mitochondria | BH3-only proteins that have homology to the BCL-2 family proteins in only a single domain, the BH3 domain |
| BIM | It promotes cell death. | The mitochondrial outer membrane (MOM) | ||
| BAD | Dephosphorylated BAD forms a heterodimer with Bcl-xL, and Bcl-2, and Bcl-xL, inactivating them and thus allowing Bax/Bak-triggered apoptosis | Free in mitochondria | ||
| BIK | It promotes Ca2 release from, activation of ER-localized Bax/ Bak. | ER | ||
| BMF | It acts as an initiator of the intrinsic apoptosis pathway. | MOM | ||
| HRK | Hrk is a critical downstream effector of the JNK dependent mitochondrial apoptotic signaling pathway. | MOM | ||
| NOXA | A selective inhibitor of MCL1 | MOM | ||
| PUMA | It promotes ER Ca2 pool depletion during thapsigargin-induced apoptosis with a Bax-dependent mechanism. | MOM |
Conclusion
Histone acetylation and deacetylation are important mechanisms to regulate gene expression. The level of HDACs is generally dysregulated in HCC, it is well accepted that epigenetic events, such as histone modification play an important role in normal biological processes and tumorigenesis and that the epigenetic status is altered during cancer initiation. Because histone deacetylation is a potentially reversible change, the epigenetic histone modification represents new opportunities for cancer management by reactivation of gene silencing. The inhibition of HDACs by GDACIs can effectively induce apoptosis and suppress cancer cell proliferation. These compounds can engage both intrinsic and extrinsic apoptotic pathways.
References
-
1.
Villota-Salazar NA, Mendoza-Mendoza A, González-Prieto JM. Epigenetics: from the past to the present. Front. Life Sci. 2016;9:347-70.
-
2.
Eccleston A, DeWitt N, Gunter C, Marte B, Nath D. Epigenetics. Nature. 2007;447:395-3.
-
3.
Feinberg AP. Editor The epigenetics of cancer etiology. Sem. Cancer Biol. 2004;14:427-32.
-
4.
Taby R, Issa JPJ. Cancer epigenetics. CA Cancer J. Clin. 2010;60:376-92. [PubMed ID: 20959400].
-
5.
Sanaei M, Kavoosi F, Mohammadi M, Khanezad M. Effect of 5-aza-2′-deoxycytidine on p16INK4a, p14ARF, p15INK4b Genes Expression, Cell Viability, and Apoptosis in PLC/PRF5 and MIA Paca-2 Cell Lines. Iran J Ped Hematol Oncol. 2019;9:219-228.
-
6.
Sanaei M, Kavoosi F, Ghasemi A. Investigation of the Effect of 5-Aza-2ʹ-Deoxycytidine on p15INK4, p16INK4, p18INK4, and p19INK4 Genes Expression, Cell Growth Inhibition, and Apoptosis Induction in Hepatocellular Carcinoma PLC/PRF/5 Cell Line. Adv. Biomed. Res. 2020;9:33. [PubMed ID: 33072645].
-
7.
Sanaei M, Kavoosi F, Nasiri S. Effect of 5-aza-2ˈ-deoxycytidine on p27Kip1, p21Cip1/Waf1/Sdi1, p57Kip2, and DNA methyltransferase 1 Genes Expression, Cell Growth Inhibition and Apoptosis Induction in Colon Cancer SW 480 and SW 948 Cell Lines. Galen Med. J. 2020;9:1899.
-
8.
Sanaei M, Kavoosi F, Hosseini F. Effect of Zebularine on p16INK4a, p14ARF, p15INK4b, and DNA Methyltransferase 1 Gene Expression, Cell Growth Inhibition, and Apoptosis Induction in Human Hepatocellular Carcinoma PLC/PRF5 and Pancreatic Cancer PA-TU-8902 Cell Lines. Iran J. Pharm. Res. 2020;19:193-202. [PubMed ID: 33841535].
-
9.
Sanaei M, Kavoosi F. Investigation of the effect of zebularine in comparison to and in combination with trichostatin A on p21Cip1/Waf1/Sdi1, p27Kip1, p57Kip2, DNA methyltransferases and histone deacetylases in colon cancer LS 180 cell line. Asian Pac. J. Cancer Prev. 2020;21:1819. [PubMed ID: 32592383].
-
10.
Sanaei M, Kavoosi F. Effect of zebularine in comparison to and in combination with trichostatin a on CIP/KIP family (p21Cip1/Waf1/Sdi1, p27Kip1, and p57Kip2), DNMTs (DNMT1, DNMT3a, and DNMT3b), class I HDACs (HDACs 1, 2, 3) and class II HDACs (HDACs 4, 5, 6) gene expression, cell growth inhibition and apoptosis induction in colon cancer LS 174T cell line. Asian Pac. J. Cancer Prev. 2020;21:2131. [PubMed ID: 32711442].
-
11.
Sanaei M, Kavoosi F. Effect of Valproic Acid on the Class I Histone Deacetylase 1, 2 and 3, Tumor Suppressor Genes p21WAF1/CIP1 and p53, and Intrinsic Mitochondrial Apoptotic Pathway, Pro-(Bax, Bak, and Bim) and anti-(Bcl-2, Bcl-xL, and Mcl-1) Apoptotic Genes Expression, Cell Viability, and Apoptosis Induction in Hepatocellular Carcinoma HepG2 Cell Line. Asian Pac. J. Cancer Prev. 2021;22:89-95.
-
12.
Sanaei M, Kavoosi F, Roustazadeh A, Shahsavani H. In-vitro effect of the histone deacetylase inhibitor valproic acid on viability and apoptosis of the PLC/PRF5 human hepatocellular carcinoma cell line. Asian Pac. J. Cancer Prev. 2018;19:2507. [PubMed ID: 30256044].
-
13.
Kavoosi F, Sanaei M. Comparative analysis of the effects of valproic acid and tamoxifen on proliferation, and apoptosis of human hepatocellular carcinoma WCH 17 celllin. Iran J. Pediatr. Hematol. Oncol. 2018;8:12-20.
-
14.
Sanaei M, Kavoosi F, Arabloo M. Effect of curcumin in comparison with trichostatin A on the reactivation of estrogen receptor alpha gene expression, cell growth inhibition and apoptosis induction in hepatocellular carcinoma Hepa 1-6 Cell lLine. Asian Pac. J. Cancer Prev. 2020;21:1045. [PubMed ID: 32334468].
-
15.
Sanaei M, Kavoosi F. Effects of 5-aza-2ˈ-deoxycytidine and valproic acid on epigenetic-modifying DNMT1 gene expression, apoptosis induction and cell viability in hepatocellular carcinoma WCH-17 cell line. Iran J. Pediatr. Hematol. Oncol. 2019;9:83-90.
-
16.
Sanaei M, Kavoosi F, Esmi Z. The effect of 5-aza-2′-deoxycytidine in combination to and in comparison with vorinostat on DNA methyltransferases, histone deacetylase 1, glutathione S-transferase 1 and suppressor of cytokine signaling 1 genes expression, cell growth inhibition and apoptotic induction in hepatocellular LCL-PI 11 cell line. Int. J. Hematol. Oncol. Stem Cell Res. 2020;14:45. [PubMed ID: 32337014].
-
17.
Chan SL, Yeo W. Targeted therapy of hepatocellular carcinoma: present and future. J. Gastroenterol. Hepatol. 2012;27:862-72. [PubMed ID: 22369685].
-
18.
Benedetti R, Conte M, Altucci L. Targeting histone deacetylases in diseases: where are we? Antioxid. Redox Signal. 2015;23:99-126.
-
19.
Lin HY, Chen CS, Lin SP, Weng JR, Chen CS. Targeting histone deacetylase in cancer therapy. Med. Res. Rev. 2006;26:397-413. [PubMed ID: 16450343].
-
20.
Barneda-Zahonero B, Parra M. Histone deacetylases and cancer. Mol. Oncol. 2012;6:579-89. [PubMed ID: 22963873].
-
21.
Zhang H, Shang YP, Chen Hy, Li J. Histone deacetylases function as novel potential therapeutic targets for cancer. Hepatol. Res. 2017;47:149-59. [PubMed ID: 27457249].
-
22.
Peart MJ, Tainton KM, Ruefli AA, Dear AE, Sedelies KA, O’Reilly LA, Waterhouse NJ, Trapani JA, Johnstone RW. Novel mechanisms of apoptosis induced by histone deacetylase inhibitors. Cancer Res. 2003;63:4460-71. [PubMed ID: 12907619].
-
23.
Shankar S, Srivastava RK. Histone deacetylase inhibitors: mechanisms and clinical significance in cancer: HDAC inhibitor-induced apoptosis. Adv. Exp. Med. Biol. 2008;5:261-98.
-
24.
Shankar S, Singh TR, Fandy TE, Luetrakul T, Ross DD, Srivastava RK. Interactive effects of histone deacetylase inhibitors and TRAIL on apoptosis in human leukemia cells: involvement of both death receptor and mitochondrial pathways. Int. J. Mol. Med. 2005;16:1125-38. [PubMed ID: 16273296].
-
25.
Kwon SH, Ahn SH, Kim YK, Bae GU, Yoon JW, Hong S, Lee HY, Lee YW, Lee HW, Han JW. Apicidin, a histone deacetylase inhibitor, induces apoptosis and Fas/Fas ligand expression in human acute promyelocytic leukemia cells. J. Biol. Chem. 2002;277:2073-80. [PubMed ID: 11698395].
-
26.
Kang MH, Reynolds CP. Bcl-2 inhibitors: targeting mitochondrial apoptotic pathways in cancer therapy. Clin. Cancer Res. 2009;15:1126-32. [PubMed ID: 19228717].
-
27.
Neumeister P, Albanese C, Balent B, Greally J, Pestell RG. Senescence and epigenetic dysregulation in cancer. Int. J. Biochem. Cell Biol. 2002;34:1475-90. [PubMed ID: 12200040].
-
28.
Wong TS, Gao W, Li ZH, Chan JYW, Ho WK. Epigenetic dysregulation in laryngeal squamous cell carcinoma. J. Oncol. 2012;5:1-10.
-
29.
Hadley M, Noonepalle S, Banik D, Villagra A. Functional analysis of HDACs in tumorigenesis. Methods Mol. Biol. 2019;6:279-307.
-
30.
Sherr CJ. Principles of tumor suppression. Cell. 2004;116:235-46. [PubMed ID: 14744434].
-
31.
Görisch SM, Wachsmuth M, Tóth KF, Lichter P, Rippe K. Histone acetylation increases chromatin accessibility. J. Cell Sci. 2005;118:5825-34. [PubMed ID: 16317046].
-
32.
Shahbazian MD, Grunstein M. Functions of site-specific histone acetylation and deacetylation. Annu. Rev. Biochem. 2007;76:75-100. [PubMed ID: 17362198].
-
33.
Firestein R, Blander G, Michan S, Oberdoerffer P, Ogino S, Campbell J, Bhimavarapu A, Luikenhuis S, de Cabo R, Fuchs C, Hahn WC. The SIRT1 deacetylase suppresses intestinal tumorigenesis and colon cancer growth. PLoS One. 2008;3:2020-7.
-
34.
Blander G, Guarente L. The Sir2 family of protein deacetylases. Annu. Rev. Biochem. 2004;73:417-35. [PubMed ID: 15189148].
-
35.
Grunstein M. Histone acetylation in chromatin structure and transcription. Nature. 1997;389:349-52. [PubMed ID: 9311776].
-
36.
Verdin E, Dequiedt F, Kasler HG. Class II histone deacetylases: versatile regulators. Trends Genet. 2003;19:286-93. [PubMed ID: 12711221].
-
37.
Sanaei M, Kavoosi F. Histone deacetylases and histone deacetylase inhibitors: molecular mechanisms of action in various cancers. Adv. Biomed. Res. 2019;8:63. [PubMed ID: 31737580].
-
38.
Au SLK, Ng IOL, Wong CM. Epigenetic dysregulation in hepatocellular carcinoma: focus on polycomb group proteins. Front. Med. 2013;7:231-41. [PubMed ID: 23620257].
-
39.
Nakamura M, Chiba T, Kanayama K, Kanzaki H, Saito T, Kusakabe Y, Kato N. Epigenetic dysregulation in hepatocellular carcinoma: an up‐to‐date review. Hepatol. Res. 2019;49:3-13. [PubMed ID: 30238570].
-
40.
Mottamal M, Zheng S, Huang TL, Wang G. Histone deacetylase inhibitors in clinical studies as templates for new anticancer agents. Molecules. 2015;20:3898-941. [PubMed ID: 25738536].
-
41.
Freese K, Seitz T, Dietrich P, Lee SM, Thasler WE, Bosserhoff A, Hellerbrand C. Histone deacetylase expressions in hepatocellular carcinoma and functional effects of histone deacetylase inhibitors on liver cancer cells in-vitro. Cancers. 2019;11:1587.
-
42.
Ding G, Liu HD, Huang Q, Liang HX, Ding ZH, Liao ZJ, Huang G. HDAC6 promotes hepatocellular carcinoma progression by inhibiting P53 transcriptional activity. FEBS letters. 2013;587:880-6. [PubMed ID: 23402884].
-
43.
Cheng CC, Liu YH, Lai YCC, Hsu YH, Chao WT, Lai YS. Hypoacetylation in association with histone 3 modulation in human hepatocellular carcinoma. in vivo. 2015;29:237-42. [PubMed ID: 25792651].
-
44.
He B, Dai L, Zhang X, Chen D, Wu J, Feng X, Zhang Y, Xie H, Zhou L, Wu J, Zheng S. The HDAC inhibitor quisinostat (JNJ-26481585) supresses hepatocellular carcinoma alone and synergistically in combination with sorafenib by G0/G1 phase arrest and apoptosis induction. Int. J. Biol. Sci. 2018;14:1845.
-
45.
Buurman R, Sandbothe M, Schlegelberger B, Skawran B. HDAC inhibition activates the apoptosome via Apaf1 upregulation in hepatocellular carcinoma. Eur. J. Med. Res. 2016;21:1-6. [PubMed ID: 26744210].
-
46.
Feng G, Dong L, Shang W, Pang X, Li J, Liu L, Wang Y. HDAC5 promotes cell proliferation in human hepatocellular carcinoma by up-regulating Six1 expression. Eur. Rev. Med. Pharmacol. Sci. 2014;18:811-6. [PubMed ID: 24706304].
-
47.
Quint K, Agaimy A, Di Fazio P, Montalbano R, Steindorf C, Jung R, Hellerbrand C, Hartmann A, Sitter H, Neureiter D, Ocker M. Clinical significance of histone deacetylases 1, 2, 3, and 7: HDAC2 is an independent predictor of survival in HCC. Virchows. Archiv. 2011;459:129-39. [PubMed ID: 21713366].
-
48.
Li G, Margueron R, Hu G, Stokes D, Wang YH, Reinberg D. Highly compacted chromatin formed in-vitro reflects the dynamics of transcription activation in vivo. Mol. cell. 2010;38:41-53. [PubMed ID: 20385088].
-
49.
Wade PA. Transcriptional control at regulatory checkpoints by histone deacetylases: molecular connections between cancer and chromatin. Hum. Mol. Genet. 2001;10:693-8. [PubMed ID: 11257101].
-
50.
Eckschlager T, Plch J, Stiborova M, Hrabeta J. Histone deacetylase inhibitors as anticancer drugs. Int. J. Mol. Sci. 2017;18:1414.
-
51.
Khan O, La Thangue NB. HDAC inhibitors in cancer biology: emerging mechanisms and clinical applications. Immunol. Cell Biol. 2012;90:85-94. [PubMed ID: 22124371].
-
52.
Lane AA, Chabner BA. Histone deacetylase inhibitors in cancer therapy. J. Clin. Oncol. 2009;27:5459-68. [PubMed ID: 19826124].
-
53.
Pasyukova EG, Vaiserman AM. HDAC inhibitors: A new promising drug class in anti-aging research. Mech. Ageing. Dev. 2017;166:6-15. [PubMed ID: 28843433].
-
54.
Maes K, Menu E, Van Valckenborgh E, Van Riet I, Vanderkerken K, De Bruyne E. Epigenetic modulating agents as a new therapeutic approach in multiple myeloma. Cancers. 2013;5:430-61. [PubMed ID: 24216985].
-
55.
Sanaei M, Kavoosi F. Effect of DNA methyltransferase in comparison to and in combination with histone deacetylase inhibitors on hepatocellular carcinoma HepG2 cell line. Asian Pac. J. Cancer Prev. 2019;20:1119. [PubMed ID: 31030484].
-
56.
Wang HG, Huang XD, Shen P, Li LR, Xue HT, Ji GZ. Anticancer effects of sodium butyrate on hepatocellular carcinoma cells in-vitro. Int. J. Mol. Med. 2013;31:967-74. [PubMed ID: 23440283].
-
57.
Joachimiak R, Kaznica A, Drewa T. Influence of sodium butyrate on hepatocellular carcinoma (hepG2) and glioblastoma (C6) cell lines in vitro. Acta Pol. Pharm. 2017;64:561-3.
-
58.
Svechnikova I, Ammerpohl O, Ekström TJ. p21waf1/Cip1 partially mediates apoptosis in hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2007;354:466-71. [PubMed ID: 17239344].
-
59.
Wang YC, Kong WZ, Xing LH, Yang X. Effects and mechanism of suberoylanilide hydroxamic acid on the proliferation and apoptosis of human hepatoma cell line Bel-7402. J. B.U.ON. 2014;19:698-704.
-
60.
Zhang HY, Zhang J, Liu KB. Experimental Study on Apoptosis Effect of Histone Deacetylase Inhibitor SAHA Enhancing Cisplatin on Hepatocellular Carcinoma. J. Yangtze Univ. (Nat. Sci. Ed.). 2011;7:13-8.
-
61.
Yang W, Zhao X, Pei F, Ji M, Ma W, Wang Y, Jiang G. Activation of the intrinsic apoptosis pathway contributes to the induction of apoptosis in hepatocellular carcinoma cells by valproic acid. Oncol. Lett. 2015;9:881-6. [PubMed ID: 25621063].
-
62.
Emanuele S, D’Anneo A, Bellavia G, Vassallo B, Lauricella M, De Blasio A, Vento R, Tesoriere G. Sodium butyrate induces apoptosis in human hepatoma cells by a mitochondria/caspase pathway, associated with degradation of β-catenin, pRb and Bcl-XL. Eur. J. Cancer. 2004;40:1441-52. [PubMed ID: 15177505].
-
63.
Ogawa K, Yasumura S, Atarashi Y, Minemura M, Miyazaki T, Iwamoto M, Higuchi K, Watanabe A. Sodium butyrate enhances Fas-mediated apoptosis of human hepatoma cells. J. Hepatol. 2004;40:278-84. [PubMed ID: 14739099].
-
64.
Herold C, Ganslmayer M, Ocker M, Hermann M, Geerts A, Hahn EG, Schuppan D. The histone-deacetylase inhibitor Trichostatin A blocks proliferation and triggers apoptotic programs in hepatoma cells. J. Hepatol. 2002;36:233-40. [PubMed ID: 11830335].
-
65.
Hao D, Deng F, Shi H, Wang H, Xiao F, Sun C, Xu Y, Li P. In vivo inhibitory effect of suberoylanilide hydroxamic acid combined with sorafenib on human hepatocellular carcinoma cells. Braz. J. Pharm. Sci. 2020;56:2175-9790.
-
66.
Zhou X, Yang XY, Popescu NC. Synergistic antineoplastic effect of DLC1 tumor suppressor protein and histone deacetylase inhibitor, suberoylanilide hydroxamic acid (SAHA), on prostate and liver cancer cells: perspectives for therapeutics. Int. J. Oncol. 2010;36:999-1005. [PubMed ID: 20198346].
-
67.
Liao B, Zhang Y, Sun Q, Jiang P. Vorinostat enhances the anticancer effect of oxaliplatin on hepatocellular carcinoma cells. Cancer med. 2018;7:196-207. [PubMed ID: 29239146].
-
68.
Gillespie S, Borrow J, Zhang XD, Hersey P. Bim plays a crucial role in synergistic induction of apoptosis by the histone deacetylase inhibitor SBHA and TRAIL in melanoma cells. Apoptosis. 2006;11:2251-65. [PubMed ID: 17051334].
-
69.
Insinga A, Monestiroli S, Ronzoni S, Gelmetti V, Marchesi F, Viale A, Altucci L, Nervi C, Minucci S, Pelicci PG. Inhibitors of histone deacetylases induce tumor-selective apoptosis through activation of the death receptor pathway. Nat. Med. 2005;11:71-6. [PubMed ID: 15619634].
-
70.
Matthews GM, Newbold A, Johnstone RW. Intrinsic and extrinsic apoptotic pathway signaling as determinants of histone deacetylase inhibitor antitumor activity. Adv. Cancer Res. 2012;116:165-97. [PubMed ID: 23088871].
-
71.
Harada H, Grant S. Apoptosis regulators. Rev. Clin. Exp. Hematol. 2003;7:117-38. [PubMed ID: 14763159].
-
72.
Hongmei Z. Extrinsic and intrinsic apoptosis signal pathway review. Apoptosis and medicine: In Tech Open. 2012;32:78-88.
-
73.
Zapata JM, Pawlowski K, Haas E, Ware CF, Godzik A, Reed JC. A diverse family of proteins containing tumor necrosis factor receptor-associated factor domains. J. Biol. Chem. 2001;276:24242-52. [PubMed ID: 11279055].
-
74.
Hockenbery D, Nuñez G, Milliman C, Schreiber RD, Korsmeyer SJ. Bcl-2 is an inner mitochondrial membrane protein that blocks programmed cell death. Nature. 1990;348:334-6. [PubMed ID: 2250705].
-
75.
Ghobrial IM, Witzig TE, Adjei AA. Targeting apoptosis pathways in cancer therapy. CA: Cancer J. Clin. 2005;55:178-94. [PubMed ID: 15890640].
-
76.
Suliman A, Lam A, Datta R, Srivastava RK. Intracellular mechanisms of TRAIL: apoptosis through mitochondrial-dependent and-independent pathways. Oncogene. 2001;20:2122-33. [PubMed ID: 11360196].
-
77.
Burz C, Berindan-Neagoe I, Balacescu O, Irimie A. Apoptosis in cancer: key molecular signaling pathways and therapy targets. Acta oncol. 2009;48:811-21. [PubMed ID: 19513886].
-
78.
Kang S, Wang S, Kuida K, Yuan J. Distinct downstream pathways of caspase-11 in regulating apoptosis and cytokine maturation during septic shock response. Cell Death Differ. 2002;9:1115-25. [PubMed ID: 12232800].
-
79.
Cory S, Adams JM. The Bcl2 family: regulators of the cellular life-or-death switch. Nature Rev. Cancer. 2002;2:647-56. [PubMed ID: 12209154].
-
80.
Scorrano L, Korsmeyer SJ. Mechanisms of cytochrome c release by proapoptotic BCL-2 family members. Biochem. Biophys. Res. Commun. 2003;304:437-44. [PubMed ID: 12729577].
-
81.
Tzifi F, Economopoulou C, Gourgiotis D, Ardavanis A, Papageorgiou S, Scorilas A. The role of BCL2 family of apoptosis regulator proteins in acute and chronic leukemias. Adv. Hematol. 2012:2012.
-
82.
Chandrasekar AP, Cummins NW, Badley AD. The role of the BCL-2 family of proteins in HIV-1 pathogenesis and persistence. Clin. Microbiol. Rev. 2019;33:1-8.
-
83.
Shamas-Din A, Kale J, Leber B, Andrews DW. Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb. Perspect. Biol. 2013;5:a008714. [PubMed ID: 23545417].
-
84.
Szegezdi E, MacDonald DC, Ní Chonghaile To, Gupta S, Samali A. Bcl-2 family on guard at the ER. Am. J. Physiol. Cell Physiol. 2009;296:C941-53. [PubMed ID: 19279228].
-
85.
Guikema JE, Amiot M, Eldering E. Exploiting the pro-apoptotic function of NOXA as a therapeutic modality in cancer. Expert Opin. Ther. 2017;21:767-79.
-
86.
Liew SH, Nguyen Q-N, Strasser A, Findlay JK, Hutt KJ. The ovarian reserve is depleted during puberty in a hormonally driven process dependent on the pro-apoptotic protein BMF. Cell Death Dis. 2017;8:e2971. [PubMed ID: 28771225].
-
87.
Anilkumar U, Prehn JH. Anti-apoptotic BCL-2 family proteins in acute neural injury. Front. Cell Neurosci. 2014;8:281. [PubMed ID: 25324720].
-
88.
Urbani A, Prosdocimi E, Carrer A, Checchetto V, Szabò I. Mitochondrial ion channels of the inner membrane and their regulation in cell death signaling. Front. Cell Dev. Biol. 2021;8:1679.