
Abstract
Spinal disk herniation can induce radicular pain through chemical irritation caused by proinflammatory and immune responses. Bone marrow mesenchymal stem cells (BMSCs) are a unique type of adult stem cell with the functions of suppressing inflammation and modulating immune responses. This study was undertaken to observe the effect of intrathecal BMSCs on the treatment of mechanical allodynia and the suppression of microglial activation in a rat noncompressive disk herniation model. The model was induced by the application of nucleus pulposus (NP) to the L5 dorsal root ganglion (DRG). The study found that the use of NP in the DRG can induce abnormal mechanical pain, increase the contents of the proinflammatory factors TNF-α and IL-1β, decrease the content of the anti-inflammatory cytokine TGF-β1 and activate microglia in the spinal dorsal horns (L5) (P < 0.05). BMSC administration could increase the mechanical withdrawal thresholds dramatically, decrease the contents of IL-1β and TNF-α, increase the content of TGF-β1 significantly (P < 0.05) and inhibit microglial activation in the bilateral spinal dorsal horn. Our results indicate that BMSC administration can reduce mechanical allodynia and downregulate the expression of proinflammatory cytokines by inhibiting microglial activation in the spinal dorsal horn in a rat noncompressive disk herniation model.
Similar content being viewed by others


Introduction
Intervertebral disk herniation may cause nerve root injury and nerve root pain, which mainly manifests as hyperalgesia, allodynia and conduction velocity reduction (Olmarker et al. 1993). The prominent manifestation of patients with disk herniation is nerve root pain, which is mainly caused by local inflammation rather than mechanical disk compression (Zai et al. 2005). In fact, inflammation caused by the nucleus pulposus (NP) can directly or indirectly sensitize the nerve roots and activate microglia in the central nervous system (CNS). Activated spinal microglia proliferate, undergo morphological changes and can release nitric oxide, oxygen-free radicals and proinflammatory factors (Hill-Felberg et al. 1999; Lendahl et al. 1990; Zai et al. 2005). The release of chemokines from immune cells and the activation of spinal microglial cells mediate nerve root pain (Abbadie 2005; Tsuda et al. 2005; Watkins and Maier 2003). Increasing evidence has shown that inflammatory cytokines, including IL-1, IL-6 and IL-8, TNF-α and the enzyme cyclooxygenase-2 (COX-2), released by activated microglia may be critical in the development of nerve root pain (Casey 2011; Cuellar et al. 2004). However, the function of glia and inflammatory cytokines in neuropathic pain remains elusive. The inhibition of microglial cell hyperactivation and excessive inflammatory cytokine release may prove to be a potential effective treatment for radicular pain.
Bone mesenchymal stem cells (BMSCs) are a promising donor resource for tissue repair and regeneration because they are multipotent and can differentiate into mesodermal lineages (Tencerova and Kassem 2016). BMSCs can also evade the identification of T cells and natural killer cells due to their lack of immunogenicity. Many articles have confirmed the therapeutic effect of BMSCs. The efficacy of BMSCs in treating traumatic brain injury and spinal cord injury was proved in animal experiments (Forostyak et al. 2013; Neirinckx et al. 2015). BMSCs transplanted into a disk can also be used in the treatment of degenerative disk disease (DDD) and have achieved good early and long-term efficacy for as long as 2 years (Pettine et al. 2016; Pettine et al. 2015). The underlying mechanism may be related to the transdifferentiation of BMSCs in place of damaged cells, the production of growth factors and their anti-inflammatory functions. All these functions are crucially important for the repair of the damaged tissue.
BMSCs can be neuroprotective in a variety of nervous system injuries and are believed to have the best potential results in pain care research models (Klass et al. 2007). In recent years, BMSCs have been used to treat neuropathic pain by systemic injection and have achieved long-lasting effects in a spared nerve injury (SNI) mouse model (Siniscalco et al. 2011). Patricia et al. found that the transfer of BMSCs directly into the ipsilateral fourth lumbar dorsal root ganglion (DRG) could modulate pain generation after sciatic nerve constriction (Musolino et al. 2007). In addition, BMSCs may have the ability to preserve the inactivated state of microglia or inhibit microglial activation, preventing activated microglia from releasing excess inflammatory cytokines and damaging the tissue (Yan et al. 2013). However, the molecular mechanisms by which BMSCs exert their effects on pain behavior are still unclear and this topic requires further study.
To the best of our knowledge, there are no studies focused on the effect of BMSCs on nerve root pain induced by intervertebral disk herniation or on activated microglial cells in neuropathic pain. The aims of the present study are to investigate the effect of BMSCs on nerve root pain and microglial activation and to elucidate the underlying mechanisms of analgesia.
Materials and methods
Animals
A total of 72 male Sprague–Dawley (SD) rats and 4 transgenic male SD rats expressing green fluorescent protein (GFP) (220–250 g) were used in this experiment. The rats were housed under standard laboratory conditions (in a temperature-controlled (21 ± 1 °C) room with a normal 12-h light/12-h dark cycle) and free access to food and water for 1 week before the initiation of the experiment.
Establishment of a rat model of noncompressive disk herniation
The rat model of noncompressive disk herniation was established as described by Kim et al. (2011). The rats were anesthetized by pentobarbital sodium (50 mg/kg, intraperitoneal injection) and a median incision approximately 1.5–2 cm long was made at the center of the line connecting the two iliac crests, 0.5–1 cm above the midpoint. The L5 left inferior articular process, L6 left superior articular process and L5 left half lamina were resected to expose the left L4 nerve root and L5 dorsal root ganglion. A longitudinal incision was made in the proximal tail of the rat. After identifying the annulus fibrosis, coccygeal NP was obtained from the rat Co2/3 intervertebral disk. Then, the NP was placed in proximity to the dorsal root ganglion (DRG) of the left L5 nerve root without compression. In addition, the wound was closed by suturing layer by layer. The surgical procedures used in sham group rats were the same, except for NP implantation.
Isolation, culture and identification of BMSCs
According to our previous BMSC extraction method (Deng et al. 2016), male SD rats (transfected with the GFP gene) were killed by CO2 asphyxiation and sterilized in 75% alcohol for 10 min. The bilateral femurs and tibias were removed aseptically. Then, the attached muscle, tendon, periosteum and other fibrous connective tissues were removed carefully. Both ends of the epiphyseal plates were cut off. Using a 20-ml syringe, the marrow cavity was rinsed with DMEM (Gibco; Thermo Fisher Scientific, USA) from the long bone end. Cells in the marrow cavity were washed in a culture dish several times until the marrow cavity became white. The cells were centrifuged and re-suspended with complete medium containing 10% FBS (Science Cell), DF-12 (GIBCO) and 1% double antibody (Gibco; Thermo Fisher Scientific, Inc). These cells were collected, seeded in 10-ml T75 flasks (Corning) and incubated at 37 °C in a 5% CO2 saturated humidity incubator. After 24 h, the culture medium was replaced and the medium was then replaced every 2 days. When the cell fusion rate reached 70–80%, the cells were passaged. Third-generation CD29 (BD FITC), CD90 (BD PE), CD45 (BD APC) and CD31 (BD PE) loci were identified by flow cytometry (FAC500, Beckman Coulter, Inc., Brea, CA, USA). All antibodies were used in a side-by-side comparison. Mixed samples were used for the side-by-side comparison. All staining procedures were in accordance with the flow antibody instructions and completed within 1 h.
After purity evaluation, passage-3 cells were transfered to a 24-well plate at a density of 1 × 105 cells in 1 ml per well for confluence. Osteogenic and adipogenic differentiation media (ScienCell, San Diego, California, USA) were mixed with the medium that was replaced every 3 days. After 3 weeks, the cells were fixed and stained with alizarin red and oil red O (Sigma-Aldrich, St. Louis, MO, USA) to examine their osteogenic and adipogenic properties, respectively. The osteogenic and adipogenic differentiation abilities of the cells were evaluated under an optical microscope (Olympus, Japan).
Experimental design and GFP-BMSC administration
Thirty-six male SD rats were randomly divided into three groups: a sham-operated group (sham), a nucleus pulposus (NP)-exposed group (NP) and a NP-exposed with BMSC group (NP + BMSCs) (n = 12). Rats received cell transplants on postoperative day 1 by lumbar puncture (LP). After successful anesthesia, LP was performed at lumbar vertebra L4, as previously described (Bakshi et al. 2006). GFP-BMSCs (106/20 μl, 20 μl) or 20 μl of normal saline (NS) was injected by a 30-gauge needle over a 2-min period and the syringe was left in place for an additional minute to prevent leakage. In the sham group, the DRG needed to be exposed without NP implantation or LP. BMSCs (106/20 μl, 20 μl) and NS (20 μl) were injected by LP in the NP + BMSCs and NP groups, respectively.
Pain behavior test
Prior to the initiation of the experiment, 36 rats were housed in an animal room for approximately 1 week for environmental adaptation. The rats were free to move 30 min before the behavioral test. The mechanical threshold or paw withdrawal threshold (PWT) was measured with von Frey filaments (North Coast Medical, Inc., Gilroy, CA) using the “up-down” method (Chaplan et al. 1994). During the whole test, the rats were kept in a clear glass case. After a brief period of tranquilization, the rat sole was mechanically stimulated with von Frey fibers of different folding strengths. When the needle was bent into a “S” shape, it was retained for 3–5 s. Retraction or withdrawal of the foot of the rat indicated a positive response and a lower level of stimulation was applied. This procedure was repeated 5 times with a 5-s interval between stimulations. The occurrence of three positive responses was considered to be a real positive until no positive response no longer appeared. The level higher than this positive response was set as the 50% PWT. When no positive reaction was observed in the rat, the upper limit of stimulation was used as the 50% PWT. The 50% threshold for mechanical paw withdrawal was calculated. If the 26 g filament stimulation did not cause a retraction reaction, the mechanical threshold was recorded as 26 g. The data were analyzed using the nonparametric method. Researchers who were blinded to the experimental subgroups and protocol performed the behavioral tests on each rat 1 day before surgery and on days 1, 3, 7, 10, 14, 21 and 28 after surgery (n = 12).
Quantitative real-time polymerase chain reaction
The rats in each group were subjected to a behavioral test 10 days after the operation. After the test, six rats per group were sacrificed by anesthesia and then perfused sufficiently with PBS. The spinal dorsal horns (L4-L6) were rapidly obtained by surgery and the obtained tissue was stored in a refrigerator at − 80 °C. According to the RNA Extraction Kit instructions, the total RNA was extracted. The A260/A280 values were determined by ultraviolet spectrophotometry. The A260/A280 ratio was between 1.8 and 2.0, which met the purity requirement. Primers were designed according to the principles of fluorescence for quantitative PCR primers. The primers were synthesized by Biomedical Engineering (Table 1, Shenggong, Shanghai, China). Reverse transcription kit instructions were used for reverse transcription and the resulting cDNA products were placed in a − 20 °C refrigerator. Amplification was performed according to the kit instructions. The results were processed by 2−△△CT.
Measurement of cytokine levels by ELISA
The rats in each group were subjected to a behavioral test 10 days after the operation. Six rats per group were sacrificed by anesthesia and then perfused sufficiently with PBS. The spinal dorsal horns (L4-L6) were rapidly obtained by surgery for ELISA analysis. The tissue was pooled and homogenized in homogenization buffer (PBS, pH 7.4, containing 1% Triton X-100, 1 mM PMSF, 10 μg/ml aprotinin and 1 μg /ml leupeptin). After centrifugation at 10,000g at 4 °C for 30 min, the supernatant was aliquoted. The supernatant was assayed by rat TNF-α, IL-1β and TGF-β1 ELISA kits. TNF-α, IL-1β and TGF-β1 levels were detected strictly according to the instructions of the ELISA test kit (R&D Systems, Minneapolis, MN, USA).
Western blotting
The protein expression of Iba-1 in the damaged spinal cord tissue 10 days after SCI was detected by Western blotting analysis with Iba-1 antibodies (1:1000, Abcam, Cambridge, UK). The total protein content in each sample was determined by the BCA method. Using a horseradish peroxidase-conjugated anti-rabbit secondary antibody (1:15,000; Abmart, Shanghai, China), a densitometry analysis of the signal intensity of the Western blot was performed using Adobe Photoshop software (Adobe, USA). The level of protein from densitometry was quantitatively analyzed with Sigma Scan Pro 5 and normalized to the GAPDH levels (n = 5).
Immunohistochemical examination
To identify and locate GFP-BMSCs and study microglial activation and the effects of BMSC administration on microglial activation in the spinal dorsal horn after disk herniation, six rats per group were euthanized 10 days after surgery after behavioral testing. Sections of the spinal cord were stained for Iba-1 to detect microglia. Intrathecal injection of GFP-BMSCs yielded green fluorescence. The rats in each group were subjected to a behavioral test and six rats per group were sacrificed by an i.p. overdose injection of pentobarbital sodium. The obtained L5 dorsal root ganglion and the corresponding spinal segment were fixed for 4 h and then dehydrated with a 20% sucrose solution for 24 h until the tissue precipitated to the bottom. The obtained spinal cord tissue was sliced to obtain frozen sections. The cross-section of the spinal cord (10 μm thick) was cut on a cryotome, blocked with 10% normal donkey serum in phosphate-buffered saline (0.01 M, pH 7.2) for 40 min and incubated overnight with a primary goat anti-Iba-1 antibody (1:500, Abcam, Cambridge, UK) at room temperature. The sections were then rinsed in PBS (5 min, 3 times) and incubated with an appropriate amount of a secondary anti-goat antibody conjugated to Cy3 (Jackson ImmunoResearch Laboratories, Inc.; 1:200) or Alexa 488 (Invitrogen; 1:200) in phosphate-buffered NS for 4 h. The cells were rinsed with PBS (5 min, 3 times) and stained with DAPI for 5 min. After rinsing with PBS, 50% PBS glycerol was used to seal the plate and the samples were observed under a fluorescence microscope (Olympus). Images were captured with a charge-coupled device camera (Fluoview, Soft Imaging Solution, Olympus) connected to the microscope and the contrast and brightness were adjusted using Photoshop CS4 (Adobe Systems, Inc.USA). To prevent fluorescence quenching, all specimens were photographed within 24 h after staining.
To quantify the Iba-1 immunointensities in the dorsal horns, microscopic images were obtained from six spinal cord sections per rat. In each section, ten images were acquired randomly. The mean gray value of each frame was obtained by using the image analysis software Image-Pro 5.0. For each studied region, a representative section from each animal was used to calculate an average exposure time and background fluorescence level, as determined by the pixel intensity of stained tissue regions that were negative for Iba-1 (Gallaher et al. 2012). The investigator was unaware of the image identities and the data were pooled for each rat and manipulated.
Statistical analysis
All results are presented as the mean ± SEM or mean ± SD. Analyses were performed by using the software SPSS 21.0. Pain behavior test, ELISA and immunohistochemical data were analyzed by a one-way analysis of variance (ANOVA) followed by Student–Newman–Keuls analyses. Statistical significance was accepted at P < 0.05.
Results
The purification and verification of BMSCs in vitro
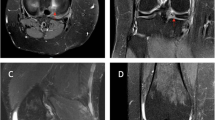
Passage-3 BMSCs were obtained using the whole marrow adherence method. Optical microscopy showed that the cell morphology was uniform, spindle-shaped, or irregularly refractive, with a high cell purity (Fig. 1a). The adherent cells grew slowly for the first 2–3 days and then proliferated rapidly, reaching confluence at 7–10 days. After five passages, the cells became large and flat and the proliferation rate decreased. Fluorescence microscopy showed that the GFP-BMSCs were labeled with green fluorescence (Fig. 1b). Under normal circumstances, the following BMSC surface markers are observed: CD29 (+), CD90 (+), CD31 (−) and CD45 (−). The CD29, CD31, CD45 and CD90 markers on the surface of the BMSCs were detected by flow cytometry. The positive rates for CD29, CD90, CD31 and CD45 were 98.8, 91.3, 0.6 and 2.4%, respectively (Fig. 2), indicating that the purity of passage-3 BMSCs was high. After a 3-week culture, the purified BMSCs exhibited osteogenic and adipogenic differentiation abilities (Fig. 1c, d), indicating that the obtained high-purity GFP-BMSCs had a good differentiation ability and were suitable for intrathecal injection.

Identification of the morphological and differentiation ability of passage-3 BMSCs extracted by whole bone marrow adherence method. a BMSCs were observed under the microscope; b BMSC-GFP cells showed green fluorescence under the fluorescence microscope; c the result of BMSCs cultured in osteogenic medium for 3 weeks: dark brown calcium nodules stained by alizarin red were observed in cells; d the result of BMSCs cultured in adipogenic medium 3 weeks: brown lipid droplets stained by oil red O were observed in cells

Detection of surface markers of passage-3 BMSCs by flow cytometry. The positive rate of CD29, CD90, CD31, CD45 on BMSCs was 98.8, 91.3, 0.6 and 2.4%, respectively
The localization of GFP-BMSCs in vivo
To identify and locate the BMSCs after intrathecal injection into the rats, we tested different spinal cord sites by immunohistochemistry 10 days after the operation. The intrathecal injection of GFP-BMSCs yielded green fluorescence and we could find some GFP-positive cells around the surface of the spinal cord approximately 2 cm above the L5 level (Fig. 3a, b). At the L5 level of the spinal cord, there were many more green cells than were observed at other sites (Fig. 1; P < 0.05). Compared with the contralateral site, the number of GFP-postive cells increased significantly at the L5 level of the ipsilateral spinal dorsal horn site (Fig. 3c, d; P < 0.05). The number of GFP-postive cells in the ipsilateral spinal ventral horn was higher than that observed on the contralateral side (Fig. 3e, f; P < 0.05). These results suggested that intrathecally injected BMSCs can locate around the surface of the spinal cord. The rat noncompressive lumbar herniation model can increase the ability of the BMSCs to migrate to the corresponding ipsilateral side of the dorsal and ventral spinal cord. BMSCs selectively migrate to the ipsilateral lumbar and dorsal spine in the noncompressive disk herniation model.

Localization of GFP-BMSCs by immunohistochemistry in the surface of the L5 level spinal cord on postoperation 10 days. a, b GFP-BMSCs showed at 2 cm above the lumbar enlargement of the spinal cord; c, d GFP-BMSCs in dorsal horn (ipsilateral side (c), contralateral side (d)); e, f GFP-BMSCs in ventral horn (ipsilateral side (e), contralateral side (f)); g the quantitative data of GFP-positive BMSCs in spinal cord.*P < 0.05 compared with control, #P < 0.05 compared with ipsilateral side
Pain behavior
There were no significant differences in PWT among the three groups before surgery (P > 0.05). From the day of surgery through 4 weeks postoperation, there was no significant decrease in PWT in the sham group (P > 0.05), while the PWT of the ipsilateral hindpaw decreased significantly in rats with lumbar disk herniation in both the NP and NP + BMSCs groups (P < 0.05). Compared with the NP group, the decrease in PWT of the ipsilateral hindpaw in the NP + BMSCs group was smaller throughout the 4-week observation period (P < 0.05, Fig. 4a). Surprisingly, the contralateral mechanical withdrawal threshold also decreased significantly between days 7 and 28 (P < 0.05, Fig. 4b) and the intrathecal injection of BMSCs attenuated mechanical allodynia in the contralateral hindpaw. Intrathecal injection of BMSCs attenuated mechanical allodynia in both the ipsilateral and contralateral hindpaws (P < 0.05) and the analgesic effect was sustained over the 4-week observation period.

Effects of BMSCs on mechanical hyperalgesia in rat models of non-oppressed disk herniation. a The mechanical withdrawal threshold for the ipsilateral hind limb between days 1 and 28 postoperatively. b The mechanical withdrawal threshold in the contralateral side hindpaw after surgery. *P < 0.05 vs sham group; #P < 0.05 vs NP group. The result was mean ± SEM of 12 rats
BMSCs reduced the contents of TNF-α and IL-1β and increased the content of TGF-β1 in the rat model
Changes in TNF-α, IL-1β and TGF-β1 mRNA and protein expression levels in the ipsilateral spinal dorsal horns (L4–L6) were evaluated by RT-PCR and ELISA, respectively (Fig. 5). It was found that in the rat noncompressive disk herniation model, the expression levels of the proinflammatory factors TNF-α and IL-1β increased significantly, while the expression level of the anti-inflammatory factor TGF-β1 was markedly decreased (P < 0.05). The expression of TNF-α and IL-1β in the NP + BMSCs group was significantly lower than that in the NP group (P < 0.05) and the expression level of TGF-β1 was significantly increased in the NP + BMSC group (P < 0.05). Intrathecal injection of BMSCs reduced the mRNA and protein levels of TNF-α and IL-1β significantly and upregulated the levels of TGF-β1 significantly.

BMSCs suppress the content of TNF-α and IL-1β and up-regulate the level of TGF-β1. The TNF-α, IL-1β and TGF-β1 protein expression levels in the ipsilateral spinal dorsal horns (L4-L6) were measured by ELISA. a, c, e The protein content of TNF-α, IL-1β and TGF-β1, respectively. The mRNA levels of TNF-α, IL-1β and TGF-β1 in ipsilateral L4-L6 spinal dorsal horns of rats were detected by PCR. b, d, f The levels of TNF-α, IL-1β, and TGF-β1 respectively. *P < 0.05 vs. sham group; #P < 0.05 vs. NP group. The result was mean ± SD of six rats
BMSCs reduced microglial activation in the spinal dorsal horn in the rat model of noncompressive disk herniation
Western blotting was used to analyze changes in Iba-1 expression in damaged spinal cord tissue harvested 10 days after SCI (Fig. 6). The protein expression levels of Iba-1 increased significantly in the NP and NP + BMSC groups compared with those in the sham group (P < 0.05) and the Iba-1 levels in the NP + BMSC group were significantly decreased compared with those of the NP group (P < 0.05). BMSC treatment downregulated the protein levels of Iba-1. Consistent with the Western blots, Iba-1-positive microglia were evenly distributed on both sides of the dorsal horn of the spinal cord of rats in the sham group by immunohistochemistry (Fig. 7a, b). Compared with the sham group, the proportion of Iba-1-positive microglia in the NP and NP + BMSC groups was significantly higher on both sides of the dorsal horn of the spinal cord. As reported in previous studies, the application of NP on the L5 DRG could upregulate Iba-1-positive microglia on both sides of the dorsal horn (Xue et al. 2014). In the NP + BMSC group, the proportion of Iba-1-positive microglia was significantly lower on both sides of the dorsal horn of the spinal cord than observed in the NP group (Fig. 7c–f). The intrathecal injection of BMSCs downregulated microglial activity in the dorsal horn of the ipsilateral and contralateral spinal cord in a rat noncompressive disk herniation model (Fig. 7g).

BMSCs suppress the content of Iba-1 protein expression levels in the spinal dorsal horns (L4-L6). *P < 0.05 vs. sham group; #P < 0.05 vs. NP group

The microglial activation in ipsilateral and contralateral sides of the dorsal horn 10 days after surgery. The red Iba-1 was the marker of the activated microglia. a, b Sham group (ipsilateral side (a), contralateral side (b)); c, d NP group (ipsilateral side (c), contralateral side (d)); e, f NP + BMSCs group (ipsilateral side (e), contralateral side (f)). g Fluorescence intensity data of Iba1 in ipsilateral and contralateral spinal cord horn. *P < 0.05 vs. sham group; #P < 0.05 vs. NP group. The result was mean ± SD of six rats
Discussion
Disk herniation is a common route of pathogenesis for clinical nerve root disease (Omarker and Myers 1998; Vucetic et al. 1999). The pathophysiology of radiculopathy is complex and incompletely understood (Childress 2017; Cohen and Hooten 2017; Eggers et al. 2016; Xue et al. 2014). The cause of nerve root disease in intervertebral disk herniation may be related to mechanical factors, inflammation, autoimmune responses and chemical insult (Kawakami et al. 1996; Kayama et al. 1996; Olmarker et al. 1993). Studies have revealed that autoimmune responses of microglia and inflammatory factors, such as TNF-α and IL-1β, are crucial for the development of nerve root pain (Ahn et al. 2002; Cuellar et al. 2004; Kallakuri et al. 2005). In the present study, we used NP on the DRG to simulate clinical disk herniation. Our study found that the pain threshold was significantly reduced when microglia in the corresponding spinal cord proliferated and became activated. The expression levels of proinflammatory cytokines released by activated microglia, such as TNF-α and IL-1β, increased significantly and the content of TGF-β1 was decreased significantly in the L4–L6 spinal dorsal horns.
The use of NP on the DGR without pressure can simulate clinical disk herniation, causing radiculopathy by increasing inflammatory cytokines and activating glial cells in the nerve roots and spinal cord (Anzai et al. 2002; Miyoshi et al. 2011; Rothman et al. 2009; Takebayashi et al. 2001). One theory holds that the NP exists as an “isolated antigen” in the human body. Once intervertebral disk protrusion occurs, the NP is exposed to the immune system, which stimulates the body to produce an autoimmune response, inducing inflammation and causing microglia (Cho et al. 2013), neutrophils, mononuclear macrophages, and other inflammatory cells to be recruited, to proliferate and to release a variety of lytic enzymes and inflammatory cytokines; this process causes nerve swelling and vacuolar degeneration, eventually leading to root neuralgia (Casey 2011; Cuellar et al. 2013; Marchand et al. 2005). Different kinds of immune cells gather in the CNS and these cells can release different factors, such as TNF-α, IL-1β, IL-6 and prostaglandins (Casey 2011).
Under physiological conditions, microglial cells in the CNS are active and monitor their surroundings (De Leo et al. 2006; Hanisch and Kettenmann 2007). Microglia that are Iba-1 (+) in the spinal cord can serve as a warning system for spinal cord and nerve root injury (Clark et al. 2007; Hubbard and Winkelstein 2005). When CNS injury occurs, microglia are activated to proliferate, undergo morphological changes and release different kinds of inflammatory cytokines (Hill-Felberg et al. 1999; Lendahl et al. 1990; Zai et al. 2005). The causes of microglial activation include simple mechanical stimulation caused by nerve compression and chemical irritation. Chemical stimulation is crucial for microglial activation (Hashizume et al. 2000). Many studies have demonstrated microglia activation in animal models of radicular pain (Cho et al. 2013; Echeverry et al. 2008; Liu et al. 1998; Rothman et al. 2009). In addition, spinal microglial activation contributes to pathological pain disease (Cao and Zhang 2008; Clark et al. 2007; Inoue 2006). Activated microglia can secrete a series of primarily inflammatory mediators and cytokines, such as TNF-α and IL-1β, which have been proposed to play important roles in the chemical pathomechanisms of radicular pain (Ahn et al. 2002; Cuellar et al. 2004; Hao et al. 2007; Leung and Cahill 2010; Sung and Wong 2007). Intrathecal injection of TNF-α and IL-1β receptor antagonists can significantly reduce nerve root pain (Kleibeuker et al. 2008; Youn et al. 2008). TGF-β1 levels decreased in rats with chronic constrictive injury, while the intrathecal injection of exogenous TGF-β1 significantly reduced the thermal pain threshold in these rats (Chen et al. 2013; Echeverry et al. 2009). In our experiment, we successfully used NP on the DRG to simulate clinical disk herniation. We found that the number of microglia and the expression levels of TNF-α and IL-1β mRNA increased significantly, while the content of TGF-β1 decreased significantly in the spinal dorsal horns (L4–L6).
BMSCs not only can be used as seed cells for differentiation into various mesodermal lineages but also have good anti-inflammatory and immunomodulatory effects (Di Nicola et al. 2002; Jiang et al. 2005; Le Blanc et al. 2004; Liu et al. 2013; Meisel et al. 2004; Sotiropoulou et al. 2006; Uccelli et al. 2008; Zanone et al. 2010). Increasing evidence indicates that BMSCs are critical for the repair of CNS damage in animal models, such as traumatic brain injury and SCI models (Baker et al. 2007; Honma et al. 2006). The effect of BMSCs on the repair of CNS damage may result from their ability to migrate, differentiate and produce growth factors (Hess et al. 2004). Furthermore, growth factors produced by BMSCs are crucial for the repair of neural damage (Jeong et al. 2012; Koh et al. 2012; Sasaki et al. 2009; Wilkins et al. 2009). Recent studies reported that BMSCs could preserve the resting phenotype of microglia and suppress the activation of microglia in vitro (Yan et al. 2013). In the present study, we demonstrated that BMSCs mediate behavioral hypersensitivity related to radicular pain by reducing inflammatory cytokine production by activated spinal microglia. On the one hand, activated microglia can effectively promote the migration and chemotaxis of exogenous BMSCs in vivo and more BMSC migration and chemotaxis promotes a stronger inhibitory effect on microglial activation. On the other hand, BMSCs can effectively reduce the contents of IL-1β and TNF-α and increase the content of TGF-β1. The release of inflammatory factors leads to the rapid activation of microglia. BMSCs can reduce the activation of microglia in the spinal cord by reducing the inflammatory response of the spinal cord. Thus, simulating the effects of endogenous anti-inflammatory and immunomodulatory mediators, such as BMSCs, may be a promising way to reduce or prevent mechanical allodynia caused by disk herniation.
One major limitation of the present study is that we only performed the experiment by using exogenous BMSCs without using intrinsic stem cell recruitment interference and immune regulation to further explore the underlying mechanism. Previous research showed that chemical stimulation of intrinsic stem cells may lead to the recovery of functions related to sensory innervation (Gallaher et al. 2014). There has been limited research about the effect of intrinsic stem cells during the process of disk herniation and whether intrinsic stem cells can be recruited to the site of injury requires further study. In addtion, the present research only concentrated on the relationship between stem cells and microglial cells without further exploring the molecular mechanism in vitro. Activated microglial cells are divided into two types: M1 and M2. There are no studies that investigated the effect of stem cells on microglial cell types and secretion. The underlying mechanisms by which BMSCs inhibit microglial activation are a question worthy of discussion and study. The conversion of M1/M2 microglia is an interesting and difficult issue in the study of microglia. In this experiment, we found that cytokines, such as IL-1β and TNF-α which are released by M1-type microglia increased significantly, while the M2-type secretory marker TGF-β1 decreased significantly in the noncompressive disk herniation model. BMSCs can decrease the contents of IL-1β and TNF-α and increase the content of TGF-β1 significantly in the dorsal horn of the spinal cord, and they can also inhibit the activation of microglia. However, the mechanism remains unclear. Therefore, further studies are needed to explore the effect of BMSCs on microglial cells of the M1/M2 types.
Conclusion
In summary, this research shows that BMSC administration can reduce mechanical allodynia and downregulate the content of proinflammatory factors and microglial activity in the spinal dorsal horn in a rat noncompressive disk herniation model, providing a new way to attenuate radicular pain. Further studies are needed to explore the effect of intrinsic stem cells during the process of disk herniation and the effect of BMSCs on M1/M2 microglia conversion and related molecular mechanisms should also be studied before BMSCs are used for clinical applications.
References
Abbadie C (2005) Chemokines, chemokine receptors and pain. Trends Immunol 26:529–534
Ahn SH, Cho YW, Ahn MW, Jang SH, Sohn YK, Kim HS (2002) mRNA expression of cytokines and chemokines in herniated lumbar intervertebral discs. Spine 27:911–917
Anzai H, Hamba M, Onda A, Konno S, Kikuchi S (2002) Epidural application of nucleus pulposus enhances nociresponses of rat dorsal horn neurons. Spine 27:E50–E55
Baker AH, Sica V, Work LM, Williams-Ignarro S, de Nigris F, Lerman LO, Casamassimi A, Lanza A, Schiano C, Rienzo M, Ignarro LJ, Napoli C (2007) Brain protection using autologous bone marrow cell, metalloproteinase inhibitors, and metabolic treatment in cerebral ischemia. Proc Natl Acad Sci U S A 104:3597–3602
Bakshi A, Barshinger AL, Swanger SA, Madhavani V, Shumsky JS, Neuhuber B, Fischer I (2006) Lumbar puncture delivery of bone marrow stromal cells in spinal cord contusion: a novel method for minimally invasive cell transplantation. J Neurotrauma 23:55–65
Cao H, Zhang YQ (2008) Spinal glial activation contributes to pathological pain states. Neurosci Biobehav Rev 32:972–983
Casey E (2011) Natural history of radiculopathy. Phys Med Rehabil Clin N Am 22:1–5
Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL (1994) Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods 53:55–63
Chen NF, Huang SY, Chen WF, Chen CH, Lu CH, Chen CL, Yang SN, Wang HM, Wen ZH (2013) TGF-beta1 attenuates spinal neuroinflammation and the excitatory amino acid system in rats with neuropathic pain. J Pain 14:1671–1685
Childress MA (2017) Spine conditions: cervical spine conditions. FP Essentials 461:11–14
Cho HK, Cho YW, Kim EH, Sluijter ME, Hwang SJ, Ahn SH (2013) Changes in pain behavior and glial activation in the spinal dorsal horn after pulsed radiofrequency current administration to the dorsal root ganglion in a rat model of lumbar disc herniation: laboratory investigation. J Neurosurg Spine 19:256–263
Clark AK, Gentry C, Bradbury EJ, McMahon SB, Malcangio M (2007) Role of spinal microglia in rat models of peripheral nerve injury and inflammation. Eur J Pain (London, England) 11:223–230
Cohen SP, Hooten WM (2017) Advances in the diagnosis and management of neck pain. BMJ 358:j3221
Cuellar JM, Montesano PX, Carstens E (2004) Role of TNF-alpha in sensitization of nociceptive dorsal horn neurons induced by application of nucleus pulposus to L5 dorsal root ganglion in rats. Pain 110:578–587
Cuellar JM, Borges PM, Cuellar VG, Yoo A, Scuderi GJ, Yeomans DC (2013) Cytokine expression in the epidural space: a model of noncompressive disc herniation-induced inflammation. Spine 38:17–23
De Leo JA, Tawfik VL, LaCroix-Fralish ML (2006) The tetrapartite synapse: path to CNS sensitization and chronic pain. Pain 122:17–21
Deng G, Wang W, Yang C, Gao R, Yang X, Ye X (2016) Shaking improves the whole bone marrow adherent method of purification. Mol Med Rep 13:3133–3138
Di Nicola M, Carlo-Stella C, Magni M, Milanesi M, Longoni PD, Matteucci P, Grisanti S, Gianni AM (2002) Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 99:3838–3843
Echeverry S, Shi XQ, Zhang J (2008) Characterization of cell proliferation in rat spinal cord following peripheral nerve injury and the relationship with neuropathic pain. Pain 135:37–47
Echeverry S, Shi XQ, Haw A, Liu H, Zhang ZW, Zhang J (2009) Transforming growth factor-beta1 impairs neuropathic pain through pleiotropic effects. Mol Pain 5:16
Eggers R, Tannemaat MR, De Winter F, Malessy MJ, Verhaagen J (2016) Clinical and neurobiological advances in promoting regeneration of the ventral root avulsion lesion. Eur J Neurosci 43:318–335
Forostyak S, Jendelova P, Sykova E (2013) The role of mesenchymal stromal cells in spinal cord injury, regenerative medicine and possible clinical applications. Biochimie 95:2257–2270
Gallaher ZR, Ryu V, Herzog T, Ritter RC, Czaja K (2012) Changes in microglial activation within the hindbrain, nodose ganglia, and the spinal cord following subdiaphragmatic vagotomy. Neurosci Lett 513:31–36
Gallaher ZR, Johnston ST, Czaja K (2014) Neural proliferation in the dorsal root ganglia of the adult rat following capsaicin-induced neuronal death. J Comp Neurol 522:3295–3307
Hanisch UK, Kettenmann H (2007) Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci 10:1387–1394
Hao S, Mata M, Glorioso JC, Fink DJ (2007) Gene transfer to interfere with TNFalpha signaling in neuropathic pain. Gene Ther 14:1010–1016
Hashizume H, DeLeo JA, Colburn RW, Weinstein JN (2000) Spinal glial activation and cytokine expression after lumbar root injury in the rat. Spine 25:1206–1217
Hess DC, Hill WD, Carroll JE, Borlongan CV (2004) Do bone marrow cells generate neurons? Arch Neurol 61:483–485
Hill-Felberg SJ, McIntosh TK, Oliver DL, Raghupathi R, Barbarese E (1999) Concurrent loss and proliferation of astrocytes following lateral fluid percussion brain injury in the adult rat. J Neurosci Res 57:271–279
Honma T, Honmou O, Iihoshi S, Harada K, Houkin K, Hamada H, Kocsis JD (2006) Intravenous infusion of immortalized human mesenchymal stem cells protects against injury in a cerebral ischemia model in adult rat. Exp Neurol 199:56–66
Hubbard RD, Winkelstein BA (2005) Transient cervical nerve root compression in the rat induces bilateral forepaw allodynia and spinal glial activation: mechanical factors in painful neck injuries. Spine 30:1924–1932
Inoue K (2006) The function of microglia through purinergic receptors: neuropathic pain and cytokine release. Pharmacol Ther 109:210–226
Jeong SR, Kwon MJ, Lee HG, Joe EH, Lee JH, Kim SS, Suh-Kim H, Kim BG (2012) Hepatocyte growth factor reduces astrocytic scar formation and promotes axonal growth beyond glial scars after spinal cord injury. Exp Neurol 233:312–322
Jiang XX, Zhang Y, Liu B, Zhang SX, Wu Y, Yu XD, Mao N (2005) Human mesenchymal stem cells inhibit differentiation and function of monocyte-derived dendritic cells. Blood 105:4120–4126
Kallakuri S, Takebayashi T, Ozaktay AC, Chen C, Yang S, Wooley PH, Cavanaugh JM (2005) The effects of epidural application of allografted nucleus pulposus in rats on cytokine expression, limb withdrawal and nerve root discharge. Eur Spine J 14:956–964
Kawakami M, Tamaki T, Weinstein JN, Hashizume H, Nishi H, Meller ST (1996) Pathomechanism of pain-related behavior produced by allografts of intervertebral disc in the rat. Spine 21:2101–2107
Kayama S, Konno S, Olmarker K, Yabuki S, Kikuchi S (1996) Incision of the anulus fibrosus induces nerve root morphologic, vascular, and functional changes. An experimental study. Spine 21:2539–2543
Kim SJ, Park SM, Cho YW, Jung YJ, Lee DG, Jang SH, Park HW, Hwang SJ, Ahn SH (2011) Changes in expression of mRNA for interleukin-8 and effects of interleukin-8 receptor inhibitor in the spinal dorsal horn in a rat model of lumbar disc herniation. Spine 36:2139–2146
Klass M, Gavrikov V, Drury D, Stewart B, Hunter S, Denson DD, Hord A, Csete M (2007) Intravenous mononuclear marrow cells reverse neuropathic pain from experimental mononeuropathy. Anesth Analg 104:944–948
Kleibeuker W, Gabay E, Kavelaars A, Zijlstra J, Wolf G, Ziv N, Yirmiya R, Shavit Y, Tal M, Heijnen CJ (2008) IL-1 beta signaling is required for mechanical allodynia induced by nerve injury and for the ensuing reduction in spinal cord neuronal GRK2. Brain Behav Immun 22:200–208
Koh SH, Baik W, Noh MY, Cho GW, Kim HY, Kim KS, Kim SH (2012) The functional deficiency of bone marrow mesenchymal stromal cells in ALS patients is proportional to disease progression rate. Exp Neurol 233:472–480
Le Blanc K, Rasmusson I, Sundberg B, Gotherstrom C, Hassan M, Uzunel M, Ringden O (2004) Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet (London, England) 363:1439–1441
Lendahl U, Zimmerman LB, McKay RD (1990) CNS stem cells express a new class of intermediate filament protein. Cell 60:585–595
Leung L, Cahill CM (2010) TNF-alpha and neuropathic pain--a review. J Neuroinflammation 7:27
Liu L, Persson JK, Svensson M, Aldskogius H (1998) Glial cell responses, complement, and clusterin in the central nervous system following dorsal root transection. Glia 23:221–238
Liu WH, Liu JJ, Wu J, Zhang LL, Liu F, Yin L, Zhang MM, Yu B (2013) Novel mechanism of inhibition of dendritic cells maturation by mesenchymal stem cells via interleukin-10 and the JAK1/STAT3 signaling pathway. PLoS One 8:e55487
Marchand F, Perretti M, McMahon SB (2005) Role of the immune system in chronic pain. Nat Rev Neurosci 6:521–532
Meisel R, Zibert A, Laryea M, Gobel U, Daubener W, Dilloo D (2004) Human bone marrow stromal cells inhibit allogeneic T-cell responses by indoleamine 2,3-dioxygenase-mediated tryptophan degradation. Blood 103:4619–4621
Miyoshi S, Sekiguchi M, Konno S, Kikuchi S, Kanaya F (2011) Increased expression of vascular endothelial growth factor protein in dorsal root ganglion exposed to nucleus pulposus on the nerve root in rats. Spine 36:E1–E6
Musolino PL, Coronel MF, Hokfelt T, Villar MJ (2007) Bone marrow stromal cells induce changes in pain behavior after sciatic nerve constriction. Neurosci Lett 418:97–101
Neirinckx V, Agirman G, Coste C, Marquet A, Dion V, Rogister B, Franzen R, Wislet S (2015) Adult bone marrow mesenchymal and neural crest stem cells are chemoattractive and accelerate motor recovery in a mouse model of spinal cord injury. Stem Cell Res Ther 6:211
Olmarker K, Rydevik B, Nordborg C (1993) Autologous nucleus pulposus induces neurophysiologic and histologic changes in porcine cauda equina nerve roots. Spine 18:1425–1432
Omarker K, Myers RR (1998) Pathogenesis of sciatic pain: role of herniated nucleus pulposus and deformation of spinal nerve root and dorsal root ganglion. Pain 78:99–105
Pettine KA, Murphy MB, Suzuki RK, Sand TT (2015) Percutaneous injection of autologous bone marrow concentrate cells significantly reduces lumbar discogenic pain through 12 months. Stem Cells (Dayton, Ohio) 33:146–156
Pettine K, Suzuki R, Sand T, Murphy M (2016) Treatment of discogenic back pain with autologous bone marrow concentrate injection with minimum two year follow-up. Int Orthop 40:135–140
Rothman SM, Guarino BB, Winkelstein BA (2009) Spinal microglial proliferation is evident in a rat model of painful disc herniation both in the presence of behavioral hypersensitivity and following minocycline treatment sufficient to attenuate allodynia. J Neurosci Res 87:2709–2717
Sasaki M, Radtke C, Tan AM, Zhao P, Hamada H, Houkin K, Honmou O, Kocsis JD (2009) BDNF-hypersecreting human mesenchymal stem cells promote functional recovery, axonal sprouting, and protection of corticospinal neurons after spinal cord injury. J Neurosci 29:14932–14941
Siniscalco D, Giordano C, Galderisi U, Luongo L, de Novellis V, Rossi F, Maione S (2011) Long-lasting effects of human mesenchymal stem cell systemic administration on pain-like behaviors, cellular, and biomolecular modifications in neuropathic mice. Front Integr Neurosci 5:79
Sotiropoulou PA, Perez SA, Gritzapis AD, Baxevanis CN, Papamichail M (2006) Interactions between human mesenchymal stem cells and natural killer cells. Stem Cells (Dayton, Ohio) 24:74–85
Sung CS, Wong CS (2007) Cellular mechanisms of neuroinflammatory pain: the role of interleukin-1beta. Acta Anaesthesiol Taiwanica : Off J Taiwan Soc Anesthesiol 45:103–109
Takebayashi T, Cavanaugh JM, Cuneyt Ozaktay A, Kallakuri S, Chen C (2001) Effect of nucleus pulposus on the neural activity of dorsal root ganglion. Spine 26:940–945
Tencerova M, Kassem M (2016) The bone marrow-derived stromal cells: commitment and regulation of Adipogenesis. Front Endocrinol 7:127
Tsuda M, Inoue K, Salter MW (2005) Neuropathic pain and spinal microglia: a big problem from molecules in "small" glia. Trends Neurosci 28:101–107
Uccelli A, Moretta L, Pistoia V (2008) Mesenchymal stem cells in health and disease. Nat Rev Immunol 8:726–736
Vucetic N, Astrand P, Guntner P, Svensson O (1999) Diagnosis and prognosis in lumbar disc herniation. Clin Orthop Relat Res:116–122
Watkins LR, Maier SF (2003) Glia: a novel drug discovery target for clinical pain. Nat Rev Drug Discov 2:973–985
Wilkins A, Kemp K, Ginty M, Hares K, Mallam E, Scolding N (2009) Human bone marrow-derived mesenchymal stem cells secrete brain-derived neurotrophic factor which promotes neuronal survival in vitro. Stem Cell Res 3:63–70
Xue F, Wei Y, Chen Y, Wang Y, Gao L (2014) A rat model for chronic spinal nerve root compression. Eur Spine J 23:435–446
Yan K, Zhang R, Sun C, Chen L, Li P, Liu Y, Peng L, Sun H, Qin K, Chen F, Huang W, Chen Y, Lv B, Du M, Zou Y, Cai Y, Qin L, Tang Y, Jiang X (2013) Bone marrow-derived mesenchymal stem cells maintain the resting phenotype of microglia and inhibit microglial activation. PLoS One 8:e84116
Youn DH, Wang H, Jeong SJ (2008) Exogenous tumor necrosis factor-alpha rapidly alters synaptic and sensory transmission in the adult rat spinal cord dorsal horn. J Neurosci Res 86:2867–2875
Zai LJ, Yoo S, Wrathall JR (2005) Increased growth factor expression and cell proliferation after contusive spinal cord injury. Brain Res 1052:147–155
Zanone MM, Favaro E, Miceli I, Grassi G, Camussi E, Caorsi C, Amoroso A, Giovarelli M, Perin PC, Camussi G (2010) Human mesenchymal stem cells modulate cellular immune response to islet antigen glutamic acid decarboxylase in type 1 diabetes. J Clin Endocrinol Metab 95:3788–3797
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant no. 81472071 and Grant no. 81301537) and the National High Technology Research and Development Program (“863” Program) of China (Grant no. 2013AA032203).
Author information
Authors and Affiliations
Contributions
Xiaodong Huang and Weiheng Wang carried out the major part of this study and wrote the manuscript. Xilin Liu helped with the animal model and behavioral testing. Yanhai Xi conducted a partial ELISA study. Jiangming Yu conducted a partial PCR study. Xiangqun Yang performed the statistical analyses and revised the manuscript. Xiaojian Ye conceived and designed the study. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Disclaimer
The content is solely the responsibility of the author.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Additional information
Dr. Xiaojian Ye is the main corresponding author of this paper.
Xiaojian Ye and Xiangqun Yang supported this project equally.
Rights and permissions
About this article

Cite this article
Huang, X., Wang, W., Liu, X. et al. Bone mesenchymal stem cells attenuate radicular pain by inhibiting microglial activation in a rat noncompressive disk herniation model. Cell Tissue Res 374, 99–110 (2018). https://doi.org/10.1007/s00441-018-2855-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-018-2855-5